C4 Plants Adaptation to High Levels of CO2 and to Drought Environments
Submitted: December 16th, 2010Reviewed: Apr 29th, 2011Published: September 22nd, 2011
DOI: 10.5772/24936
IntechOpen Downloads
26,435
Full Chapter Downloads on intechopen.com
Altmetric score
Overall attention for this chapters
María Valeria Lara*
Centro de Estudios Fotosintéticos y Bioquímicos (CEFOBI) – Facultad de Ciencias Bioquímicas y Farmacéuticas, Universidad Nacional de Rosario, Argentina
Carlos Santiago Andreo
Centro de Estudios Fotosintéticos y Bioquímicos (CEFOBI) – Facultad de Ciencias Bioquímicas y Farmacéuticas, Universidad Nacional de Rosario, Argentina
*Address all correspondence to:
one. Introduction
one.i. General features of the C4 bicycle
All plants employ the Photosynthetic Carbon Reduction (PCR or Calvin-Benson) cycle for COtwo fixation in which Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) catalyzes the commencement stride producing a 3-carbon compound, phosphoglycerate (three-PGA). For this reason this procedure is referred to as the Cthree cycle. Plants utilizing this pathway are often named as C3 species. A major problem with the C3 cycle is that the enzyme Rubisco catalyzes two competing reactions: carboxylation and oxygenation (Portis & Parry, 2007). The oxygenation reaction directs the flow of carbon through the photorespiratory pathway, and this can result in losses of between 25% and thirty% of the carbon stock-still. Environmental variables such as high temperature and drought can result in an increase in the oxygenase reaction. Therefore, reducing the Rubisco oxygenase reaction has the potential to increment carbon assimilation significantly and would stand for a stride change in photosynthesis (up to 100% depending on temperature; Long et al., 2006).
The Civ photosynthesis is an adaptation of the C3 pathway that overcomes the limitation of the photorespiration, improving photosynthetic efficiency and minimizing the h2o loss in hot, dry environments (Edwards & Walker, 1983). Generally, Cfour species originate from warmer climates than C3 species (Sage & Monson, 1999). Most Cfour plants are native to the tropics and warm temperate zones with high light intensity and high temperature. Nether these weather, C4 plants showroom college photosynthetic and growth rates due to gains in the water, carbon and nitrogen efficiency uses. Indeed, the highest known productivity in natural vegetation is for a C4 perennial grass in the primal Amazon, which achieves a net production of 100 t (dry matter) ha-one year-1 (Piedade et al., 1991, Long, 1999). Some of the world´s near productive crops and pasture, such as maize (Zea mays), carbohydrate cane (Saccharum officinarum), sorghum (Sorghum bicolor), amaranth, paspalums (Paspalum notatumand P. urvillei),bermudagrass (Cynodon dactylon), blue grama (Bouteloua gracilis)and rhodes grass (Chloris gayana)are Cfour plants. In addition, the most troublesome weeds similar nutgrass, crabgrass and undiscriminating, are also C4 species. Although C4 plants stand for only a small portion of the world´due south constitute species, accounting for only 3 % of the vascular plants, they contribute well-nigh 20% to the global principal productivity because of highly productive C4-grass-lands (Ehleringer et al., 1997). Approximately half of the ˜ten,000 grass and sedge species have Civ photosynthesis, but fewer than 2,000 of the dicotyledonous species exhibit C4 photosynthesis. Given their disproportionate influence on global productivity, C4 plants take attracted much attention by the ecophysiological and ecosystem communities (Sage & Monson, 1999).
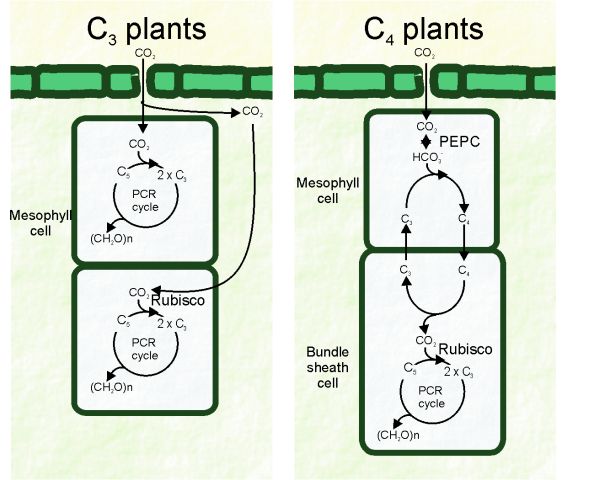
In C4 plants, the photorespiration is suppressed by elevating the CO2 concentration at the site of Rubisco though suppressing the oxygenase action of the enzyme. This is accomplished by a biochemical COii pump and relies on a spatial separation of the COtwo fixation and assimilation. In full general, these species take a particular beefcake (Kranz beefcake), where mesophyll and bundle sheath cells cooperate to fix CO2 (Effigy 1). Differentiation of these two jail cell types is essential for the operation of C4 photosynthesis, although special cases for the performance of the C4 cycle within only ane type of photosynthetic cell accept been found (Edwards et al., 2004, Lara et al., 2002, Lara & Andreo, 2005).
Basically, carboxylation of phosphoenolpyruvate (PEP) by the phosphoenolpyruvate carboxylase (PEP-carboxylase) produces four-carbon organic acids in the cytosol of mesophyll cells. This so-called C4 compounds are transported to the packet sheath cells and decarboxylated to yield COtwo which is assimilated by Rubisco in the Photosynthetic Carbon Reduction (PCR) bike (Hatch, 1987). The decarboxylation reaction besides produces iii-carbon organic acids (C3) that return to the mesophyll cells to regenerate PEP in a reaction catalyzed past the enzyme pyruvate orthophosphate dikinase (PPDK). This process called
Hatch-Slack pathway, after the first scientists that postulated the metabolic pathway. However, they used the proper noun Civdicarboxylic acrid pathway of photosynthesis.Due to current use, the proper name has been shortened to C4photosynthesis, C4pathway, C4syndromeor Civmetabolism. The plants that perform this type of photosynthesis are then called C4plants.
This general scheme is common among the C4 species; however, in that location are variations to this basic pathway that include various decarboxylation enzymes every bit well as unlike transported metabolites. Thus, the decarboxylation procedure occurs in 3 various means, mainly using one of the following enzymes: NADP-malic enzyme (NADP-ME), NAD-malic enzyme (NAD-ME) or phosphoenolpyruvate carboxykinase (PEP-CK). Therefore, C4 plants have been traditionally grouped into iii biochemical subtypes depending on the major decarboxylase used (C4-NADP-ME subtype; Cfour-NAD-ME subtype or C4-PEP-CK subtype). Each C4 subgroup possesses particular structural features, biochemistry and physiology, and also differences in the mechanism used to regenerate phosphoenolpyruvate (PEP), the substrate of PEP-carboxylase in mesophyll cells. Yet, it is now becoming apparent that, in several cases, more than than one decarboxylase operates at the aforementioned time (Drincovich et al., 2011).
one.two. Ciii vs C4 species
Cfour species have evolved in a high CO2 environment. This increases both their nitrogen and water use efficiency compared to Ciii species. Civ plants have greater rates of COtwo assimilation than Cthree species for a given leaf nitrogen when both parameters are expressed either on a mass or an area basis (Ghannoum et al., 2011). Although the range in leafage nitrogen content per unit areas is less in C4 compared to Ciii plants, the range in leaf nitrogen concentration per unit dry out mass is similar for both C4 and Ciii species. Even though leaf nitrogen is invested into photosynthetic components into the aforementioned fraction in both Ciii and C4 species, Civ plants allocate less nitrogen to Rubisco protein and more than to other soluble protein and thylakoids components. In C3 plants, the photosynthetic enzyme Rubisco accounts for upwards to xxx% of the leaf nitrogen content (Lawlor et al., 1989), but accounts for only four–21% of leaf nitrogen in C4 species (Evans & von Caemmerer, 2000, Sage et al., 1987). The lower nitrogen requirement of C4 plants results from their CO2-concentrating machinery, which raises the bundle sheath CO2 concentration, saturating Rubisco in normal air and almost eliminating photorespiration. Without this mechanism, Rubisco in the C3 photosynthetic pathway operates at but 25% of its capacity (Sage et al., 1987) and loses ca. 25% of fixed carbon to photorespiration (Ludwig & Canvin, 1971). To attain comparable photosynthetic rates to those in C4 plants, C3 leaves must therefore invest more than heavily in Rubisco and accept a greater nitrogen requirement. Because the Rubisco specificity for COtwo decreases with increasing temperature (Long, 1991), this departure between the Ciii and Cfour photosynthetic nitrogen-apply efficiency is greatest at high temperatures (Long, 1999). The high photosynthetic nitrogen-employ efficiency of C4 plants is partially offset by the nitrogen-requirement for COii-concentrating machinery enzymes, just the high maximum catalytic rate of PEP-carboxylase means that these account for just ca. five% of leaf nitrogen (Long, 1999). Improved leaf and establish water apply efficiency in Cfour plants is due to both college photosynthetic rates per unit leafage expanse and lower stomatal conductance, with the greater CO2 assimilation contributing to a major extent (Ghannoum et al., 2011).
The advantages of greater nitrogen apply efficiency and h2o use efficiency of C4 relative to C3 photosynthesis are fully realized at high light and temperature, where oxygenase reaction of Rubisco is profoundly increased. It is worth noting, although in C4 plants energy loss due to photorespiration is eliminated, and additional energy is required to operate the Civ bike (2 ATPs per CO2 assimilated). In dim light, when photosynthesis is linearly dependent on the radiative flux, the rate of CO2 assimilation depends entirely on the free energy requirements of carbon assimilation (Long, 1999). The additional ATP required for assimilation of i CO2 in Cfour photosynthesis, compared with Ciii photosynthesis, increases the free energy requirement in C4 plants (Hatch, 1987). However, when the temperature of a Cthree leaf exceeds ca. 25 ºC, the corporeality of light energy diverted into photorespiratory metabolism in C3 photosynthesis exceeds the additional free energy required for COtwo assimilation in C4 photosynthesis (Hatch, 1992, Long, 1999). This is the reason why at temperatures below ca. 25–28 ºC, Cfour photosynthesis is less efficient than C3 photosynthesis under light-limiting conditions. It is interesting to note, that while global distribution of C4 grasses is positively correlated with growing flavour temperature, the geographic distribution of the unlike Cfour subtypes is strongly correlated with rainfall (Ghannoum et al., 2011).
On the contrary, C4 plants are rare to absent in cold environments. Although at that place are examples of plants with C4 metabolisms that evidence cold adaptation, they still require warm periods during the day in gild to exist in cold habitats (Sage et al., 2011). In consequence, C4 species are poorly competitive against C3 plants in common cold climates (Sage & McKown, 2006, Sage & Pearce, 2000). The mechanisms explaining the lower performance of Civ plants nether cold conditions have not been antiseptic (Sage et al., 2011). Amongst early plausible explanations were the depression quantum yield of the Cfour relative to the C3 pathway (Ehleringer et al., 1997), and enzyme lability in the C4 bicycle, most notably around PEP metabolism (PEP-carboxylase and pyruvate orthophosphate dikinase) (Matsuba et al., 1997). Both hypothesis are insufficient since maximum breakthrough yield differences do not relate to conditions under which the vast majority of daily carbon is assimilated and in that location cold-adapted C4 species that have cold stabled forms of PEP-carboxylase and pyruvate orthophosphate dikinase, and synthesize sufficient quantity to overcome any short term limitation (Du et al., 1999, Hamel & Simon, 2000, Sage et al., 2011). The current hypothesis is that C4 photosynthesis is express past Rubisco capacity at low temperatures. Fifty-fifty in cold-tolerant C4 species, Rubisco capacity becomes limiting at depression temperature and imposes a ceiling on photosynthetic rate below20 ºC (Kubien et al., 2003, Pittermann & Sage, 2000, Sage,2002).
Advertisement
2. Climate change
According to the Intergovernmental Panel on Climate Change (IPCC), the current atmospheric CO2 level of 384 μmol l-1 (800 Gt) is predicted to ascent to one thousand Gt by the year 2050. Only this time humans are the drivers of these changes and non glacial-interglacial cycles. Human being-acquired increases in atmospheric COtwo concentration are thought to be largely responsible for recent increases in global hateful surface temperatures and are projected to increase by i.4 to over 5 ºC past 2100 (Intergovernmental Panel on Climate Change, 2001, 2007). Increment in global average temperatures would farther issue in drastic shifts in the almanac atmospheric precipitation with a 20% reduction per year, and virtually 20% loss in soil moisture (Schiermeier, 2008). Regarding plants, college atmospheric CO2 levels tend to reduce stomatal conductance and transpiration, thereby lowering latent estrus loss and causing higher leaf temperatures (Bernacchi et al., 2007). Thus, in the future, plants will likely experience increases in acute heat and drought stress, which can touch ecosystem productivity (Cias et al., 2005) and biodiversity (Thomas et al., 2004). The sensitivity of photosynthesis to each of the environmental variables including high temperature, low h2o availability, vapor force per unit area deficit and soil salinity, associated with the inevitable rise in atmospheric CO2, has not been well documented in assessing constitute responses to the new changing surroundings (Reddy et al., 2010). How plant growth responds to the rising CO2 concentration will not only affect ecosystem productivity in the hereafter, but also the magnitude of C sequestration by plants and, consequently, the charge per unit of COtwo increment in the temper. C4 plants are straight affected by all major global alter parameters, frequently in a manner that is distinct from that of C3 plants. In the present chapter, we volition focus on the effect of increased CO2, and its relation to temperature and drought, on C4 plants. Understanding how plants have and volition respond to the rapid change in CO2 concentration, together with developing cognition about their chapters to adapt, is an essential initial stride in agreement the full impact that the multiple interacting factors of global change (e.g. drought, temperature, ozone) will have on terrestrial ecosystems. These ecosystems produce services upon which nosotros are dependent for food, fuel, fiber, clean air, and fresh h2o (Leakey et al., 2009).
Advertisement
iii. The CO2 response
In theory, increases in atmospheric levels of COii above current levels can increase photosynthesis by decreasing photorespiration (fixation of O2 rather than CO2 by Rubisco), which increases with temperature and is higher in C3 than Civ and crassulacean acrid metabolism (CAM) plants (Sage & Monson, 1999). In improver, ascent COii generally stimulates C3 photosynthesis more than C4. Doubling of the current ambient CO2 concentration stimulated the growth of C4 plants to the melody of 10–20% whereas that in Ciii plants was about twoscore–45% (Ghannoum et al., 2000).
C3 photosynthesis is known to operate at less than optimal CO2 levels and tin evidence dramatic increase in carbon assimilation, growth and yields. Equally Rubisco is substrate-express by the electric current atmospheric COii levels, this enzyme has the potential to reply to increases in CO2 concentration; and have a metabolic control to alter the COtwo flux during carbon assimilation (Bernacchi et al., 2003, Long et al., 2004). On the contrary, photosynthetic carbon assimilation in the Cfour species is saturated or virtually CO2-saturated a low ambient pCOtwo. The reason is that PEP-carboxylase utilizes HCO3- equally substrate rather than CO2; in outcome, the enzyme is insensitive to changes in the ratio of CO2: O2 due to lack of binding of O2 to the catalytic site of PEP-carboxylase. Therefore, if plants were grown under elevated COtwo, carbon fixation would be piffling affected. This assumption that the inherent COtwo concentrating mechanism in C4 plants renders these plants insensitive to elevated CO2 atmosphere is reflected in the lack of interest that information technology has been attributed to the study of the C4 plants response to elevated COtwo levels. To show this, Reddy et al. (2010) performed an exhaustive fifteen year- literature survey on the influence of elevated COtwo among sure C3, Cfour and CAM species. The authors provided information for forty C3 plants and for only two C4 species and iii CAM plants. Most of the C3 plants presented a significant positive response to photosynthetic acclimation, Sorghum and Panicum (C4 plants) exhibited negative response, whereas Ananas, Agave and Kalanchoe (CAM plants) showed positive responses to increased CO2 concentration during growth. In view of this survey, it is then evident, that responses to elevated CO2 accept been little investigated in Cfour species. Moreover, conflicting reports on institute responses to elevated CO2, and several such differential photosynthetic responses, could be attributed to differences in experimental technologies, plant species used for the experiments, age of the plant as well as duration of the treatment (Sage, 2002). Nevertheless, C4 species notwithstanding showroom positive responses (Fig. 2), particularly at elevated temperature and arid conditions where they are currently common and under food-express situations every bit well (Ghannoum et al., 2000, Sage & Kubien, 2003). High COii aggravates nitrogen limitations and in doing so may favor Civ species, which take greater photosynthetic nitrogen use efficiency (Sage & Kubien, 2003). On the other hand, elevated COii can as well increase water utilise efficiency, in part by decreasing stomatal conductance and transpiration (Ainsworth et al., 2002). The irradiance is also a paramount factor; enhanced photosynthesis under elevated CO2 conditions was observed in C4 plants grown nether loftier irradiance, while in that location was not much response when grown nether low irradiance (Ghannoum et al., 2000).
Differences in the conductance of the bundle sheath cells to CO2 (varying with the decarboxylating subtype and as well associated with changes in the ratio of Rubisco:PEP-carboxylase activity) were proposed to be responsible for different rates of CO2 leakage (Brown & Byrd, 1993, Ehleringer & Pearcy, 1983, Hattersley, 1982, Saliendra et al., 1996). Nevertheless, further studies showed that the stimulation of foliage photosynthesis at elevated CO2 was not associated with COii leak rates from the bundle sheath or with changes in the ratio of activities of PEP-carboxylase to Rubisco (Ziska et al., 1999).
Some other attribute of plant metabolism which may vary under exposure to increased CO2 is the respiration. As highlighted by Reddy and colleagues (2010) in Civ plants little is known about the impact of elevated COii on the respiratory rates, which are reduced in C3 species and thus, probably contributing to increase biomass yield.
Neither Cthree nor Civ species show acclimation responses that are directly linked to CO2 level. Instead, the CO2 consequence on the photosynthetic biochemistry is largely mediated by carbohydrate accumulation in leaves under atmospheric condition where carbon sinks in the plant are also experiencing high carbon supply (Sage & McKown, 2006). The effectiveness with which increases in COtwo tin can be translated into growth benefits is depending in the sink-source balance and is affected by various plant and ecology factors. Depending on the growing weather condition, these changes may or not conduct to increases in leaf area (Ghannoum et al., 2001, Leakey et al., 2006, Morison & Lawlor, 1999). For plants grown under optimal growth weather and elevated COii, photosynthetic rates can be more than 50% higher than for plants grown under normal CO2 concentrations. This reduces to 40% higher for plants grown under the average of optimal and suboptimal conditions, and over the form of a full day, boilerplate photosynthetic enhancements under elevated COtwo are estimated to exist most 30%. The 30% enhancement in photosynthesis is reported to increment relative growth rate by only nigh 10%. This discrepancy is probably due to enhanced carbohydrate availability exceeding many plants' power to fully utilize it due to nutrient or inherent internal growth limitations. Consequently, growth responses to elevated CO2 increase with a plant'due south sink capacity and nutrient status (Kirschbaum, 2010).
3.1. Responses to increased CO2 levels are dependent on other environmental factors
3.1.ane. Increased CO2 and drought
Global circulation models accept predicted that, together with increases in the COii concentration, in the future some regions will have increases in the frequency and severity of droughts.
Leaky et al. (2009) proposed that the potential for increased growth and yield of C4 plants at elevated CO2 concentrations relays on the decrease in water use and reduction of drought stress, and non by a direct upshot of increased photosynthesis. In this respect, some Cfour plants
Effigy two.
Summary of the main factors involved in the response of plants to elevated COtwo
grown under Complimentary-Air Carbon dioxide Enrichment (Face up) exhibited increased photosynthetic rates only during drought or under the conditions of atmospheric vapor pressure deficits (Cousins, et al., 2002, Leakey et al., 2009). Elevated CO2 reduced midday stomatal conductance of Face up-grown sorghum by 32% with irrigation and by 37% under drought stress (Wall et al., 2001). The effect of elevated COii concentration on whole plant h2o apply was smaller, but even so meaning (Conley et al., 2001). It is worth mentioning, that this indirect mechanism of enhanced carbon uptake by elevated CO2 concentration is not unique to C4 plants. Decreased stomatal conductance at elevated concentration of CO2 in a C3 soybean awning likewise led to a significant reduction in awning evapo-transpiration (Bernacchi et al., 2007). Therefore, interactive effects of CO2 and water availability may alter the relative performance of C3 and Cfour species. At stated before, at current CO2 levels, C4 species (particularly dicots) generally require less water than Ciii because of the higher CO2 uptakes rates and greater stomatal resistance to water loss (Ehleringer et al., 1997). Under conditions of drought and elevated CO2, based on comparative studies using model C3 and C4 plants, Ward et al. (1999) postulated that Cthree species would be more competitive than Civ species as results of decreased water loss through transpirations and higher CO2 rates that would decrease the relative advantage of Civ plants nether drought conditions.
three.ane.2. Increased COii and temperature
Global increases in temperature and CO2 may have interactive effects on photosynthesis. On one hand, negative effects of heat stress on plants are well known, since photosynthesis is idea to be among the nearly thermosensitive aspects of institute function. Both the low-cal (electron transport) and dark (Calvin cycle) reactions of photosynthesis take thermolabile components, especially photosystem Ii (PSII) in the light reactions (Berry & Björkman, 1980, Heckathorn et al., 1998, 2002, Santarius 1975, Weis & Berry, 1988) and Rubisco activase in the Calvin cycle (Crafts-Brandner & Salvucci, 2002). Therefore, limiting processes decision-making photosynthesis at elevated temperature could be either failing capacity of electron send to regenerate ribulose-ane,5-bisphosphate, or reductions in the chapters of Rubisco activase to maintain Rubisco in an agile configuration (Sage et al., 2008).
Since, studies examining the effects of elevated COtwo and increased growth temperature (typically 3–5 ◦C) had yield positive (Faria et al., 1996, 1999, Ferris et al.,1998, Huxman et al., 1998, Taub et al., 2000), negative (Bassow et al., 1994, Roden & Ball, 1996), and no effects (Coleman et al., 1991) on photosynthetic and institute tolerance to acute rut stress. Once more, growing conditions and type of carbon assimilation pathways are need to exist discriminated. Full general effects of elevated CO2 on photosynthetic heat tolerance were recently investigated in a comparative study including C3 and C4 species and they can be summarized every bit follows: (i) in C3 species, elevated CO2 typically increases oestrus tolerance of photosynthesis, except for plants grown at supra-optimal growing temperature, and so elevated CO2 may provide no benefit or even decrease photosynthesis; (ii) in C4 species, elevated CO2 frequently decreases photosynthetic thermotolerance, at near-optimal growing temperature likewise as supra-optimal growing temperature (Wang et al. 2008; Hamilton et al., 2008). Although both C3 and Cfour plants experience reductions of similar magnitude in stomatal conductance with increasing COtwo (e.g., 20%–l% with a doubling of CO2) (Sage, 1994; Reich et al., 2001; Wang et al., 2008), the lower stomatal conductance of C4 plants at any given COtwo level means lower average transpiration and higher leaf temperatures in C4 plants, which may increase heat related damage in Cfour plants compared with C3 plants in the same habitat. On the other manus, elevated COtwo increases leaf size (Morison & Lawlor, 1999), and this should increment leaf temperatures during heat stress more in C3 than C4 species, given the greater average stimulation of growth in elevated CO2 in Ciii species (Poorter & Navas, 2003).
3.1.3. Other considerations
Finally, to accept a deeply understanding of the performance of C4 plants under increased CO2 atmospheric condition other factors besides water availability, soil nutrition and temperature, should be considered. I aspect to be included in the analysis should exist pests and diseases.
Changes in the ratio of CO2/O2 in the atmosphere affects establish metabolism in ways that ultimately influence the quality of leaves every bit a nutrient resource for animals. To herbivores, the decreased leaf protein contents and increased carbon/nitrogen ratios common to all leaves nether elevated atmospheric carbon dioxide imply a reduction in nutrient quality. Stiling and Cornelissen (2007) analyzed found-herbivore interactions using Ciii species and plant that plants grown under elevated CO2 usually had lower nutrient concentrations, which reduced the growth rate of herbivores feeding on that found textile. Contrasting C4 and C3 species, C4 grasses are a less nutritious food resource than Ciii grasses, both in terms of reduced poly peptide content and increased carbon/nitrogen ratios. The abundance of C3 and Civ plants (particularly grasses) are affected by atmospheric carbon dioxide. There is an indication that every bit C4-dominated ecosystems expanded vi–viii Ma b.p., there were significant species-level changes in mammalian grazers. Today in that location is bear witness that mammalian herbivores differ in their preference for C3 versus C4 food resources, although the factors contributing to these patterns are not clear. Elevated carbon dioxide levels volition likely alter nutrient quality to grazers both in terms of fine-scale (protein content, carbon/nitrogen ratio) and coarse-calibration (C3 versus Cfour) changes (Ehleringer et al., 2002).
Regarding constitute-constitute interactions using C3 species, Wang (2007) showed that the growth response of mixed-species communities to elevated CO2 was less than the response of single-species populations. In addition, the relative importance of these and other factors should be established for C4 species grown under elevated COtwo.
Advertizing
four. Conclusion
Cfour plants are directly affected past all major global change parameters, often in a fashion that is distinct from that of Ciii plants. Although an ongoing endeavor has been dedicated to the study of the response of C4 plants to CO2 enrichment, the literature regarding the response of C4 plants is yet under-represented when comparison to that of C3 species. An understanding of Cfour plants responses to ambient variables such as temperature, CO2, nutrients and water is essential for predictions of how agricultural and wild C4 populations will respond to climate variations such as those predicted to occur with global climate change (Intergovernmental Panel on Climate change, IPCC, 2001).
Advertizing
Acknowledgments
This piece of work was funded past a grant from Agencia Nacional de Promoción Científica y Tecnológica (PICT Nº 2008-2164) and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, PIP Nº0679). CSA and MVL and are members of the Researcher Career of CONICET.
References
1.AinsworthEastward. A.DaveyP. A.BernacchiC. J.DermodyO. C.HeatonDue east. A.MooreD. J.MorganP. B.NaiduS. L.RaH. Due south. Y.ZhuX. G.et al.2002A metaanalysis of elevated [COii] furnishings on soybean (Glycine max) physiology, growth and yield. Global Modify Biology8695-709.
two.BassowSouthward. Fifty.Mc ConnaughayOne thousand. D. M.BazzazF. A.1994The response of temperate tree seedlings grown in elevated COii to extreme temperature events.Ecological Applications4593-603.
3.BernacchiC. J.PimentelC.LongSouth. P.2003In vivo temperature response functions of parameters required to model RuBP-express photosynthesis.Institute Prison cell and Environment2614191430.
4.BernacchiC. J.KimballB. A.QuarlesD. R.LongS. P.OrtD. R.2007Decreases in stomatal conductance of soybean under open air elevation of CO2 are closely coupled with decreases in ecosystem evapotranspiration,Constitute Physiology14313444.
5.BerryJ. A.BjörkmanO.1980Photosynthetic response and accommodation to temperature in higher plants.Annual Reviews in Plant Physiology31491543.
half dozen.BrownR. H.ByrdChiliad. T.1993Estimation of Bundle Sheath cell conductance in C4 species and O2 insensitivity of photosynthesis.Plant Physiology.11831188-.
7.CiaisP. H. .ReichsteinM. .ViovyNorth. .GranierA. .OgeeJ. .AllardV.et al.2005Europe-wide reduction in primary productivity caused by the heat and drought in 2003.Nature437529-533.
8.ColemanJ. S.RochefortL.BazzazF. A.WoodwardF. I.1991Atmospheric CO2, institute nitrogen status and the susceptibility of plants to acute heat stress.Plant Cell and Surround14667-674.
x.CousinsA. B.et al.2002Photosystem Ii energy use, non-photochemical quenching and the xanthophyll cycle in Sorghum bicolor grown under drought and free-air COtwo enrichment (FACE) conditions. Institute Prison cell and Environs251551-1559.
11.Crafts-BrandnerS. J.SalvucciM. Eastward.2002Sensitivity of photosynthesis in a Civ plant, maize, to heat stress.Plant Physiology1291773-1780.
12.DrincovichM. F.LaraG. V.MaurinoV. Thousand.AndreoC. Southward.2011C4 Decarboxylases: Dissimilar Solutions for the Same Biochemical Problem, the Provision of CO2 to Rubisco in the Package Sheath Cells. In: Raghavendra, A.S. & Sage, R.S. (Eds.)C4 Photosynthesis and Related COtwo Concentrating Mechanisms,Springer Scientific discipline+Business Media B.V., Dordrecht, The Netherlands,277300.
thirteen.DuY. N.Olfactory organA.WasanoGrand.1999Thermal feature of C4 photosynthetic enzymes from leaves of iii sugarcane species differing in cold sensitivity.Plant and Jail cell Physiology40298-304.
14.EdwardsOne thousand. East.WalkerD. A. .Eds1983C3, C4: mechanisms, and cellular and environmental regulation of photosynthesis,Blackwell Scientific, Oxford.
15.EdwardsK. East.FranceschiV. R.VoznesenskayaE. V.2004Single-prison cell C4 photosynthesis versus the dual-cell (Kranz) paradigm.Annual Review in Constitute Biology5517396.
xvi.EhleringerJ.PearcyR. Due west.1983Variation in quantum yield fro CO2 uptake among C3 and C4 plants.Plant Physiology73555-559.
17.EhleringerJ. R.CerlingT. Eastward.DearingM. D.2002Atmospheric COtwo equally a Global Modify Commuter Influencing Plant-Animal Interactions.Integrative and Computational Biology42424-430.
eighteen.EhleringerJ. R.CerlingT. E.HellikerB. R.1997C4 photosynthesis, atmospheric COii and climate.Oecologia112285-299.
19.EvansJ. R.vonCaemmerer. Southward.2000Would Civ rice produce more than biomass than Cthree rice? In: Hardy, B. (Ed.)Redesigning rice photosynthesis to increment yield,International Rice Inquiry Constitute and Elsevier Science BV, Amsterdam, Netherlands,5371.
twenty.FariaT.VazChiliad.SchwanzP.PoleA.PereiraJ. S.ChavesM. M.1999Responses of photosynthetic and defense systems to high temperature stress in Quercus suber L. seedlings grown under elevated CO2. Plant Biologyane365-371.
21.FariaT.WilkinsD.BesfordR. T.VazM.PereiraJ. S.ChavesYard. Grand.1996Growth at elevated CO2 leads to down-regulation of photosynthesis and altered response to high temperature in Quercus suber L. seedlings. Journal of Experimental Botany471755-1761.
22.FerrisR.WheelerT. R.HadleyP.EllisR. H.1998Recovery of photosynthesis after environmental stress in soybean grown under elevated CO2.Crop Scientific discipline38948-954.
23.GhannoumO.EvansJ. R.vonCaemmerer. Southward.2011Nitrogen and water utilize efficiency of C4 plants. In: Raghavendra, A.Southward. & Sage, R.S. (Eds.)C4 Photosynthesis and Related CO2 Concentrating Mechanisms,Springer Science+Business Media B.V., Dordrecht, Holland,129146.
24.GhannoumO.vonCaemmerer. S.ConroyJ. P.2001Plant water use efficiency of 17 NAD-ME and NADP-ME C4 grasses at ambient and elevated CO2 partial pressure.Australian Journal of Establish Physiology281207-1217.
25.GhannoumO.vonCaemmerer. S.ZiskaL. H.ConroyJ. P.2000The growth response of C4 plants to ascent atmospheric COtwo partial pressure level: a reassessment.Plant, Cell and Surround23931-942.
26.HamelN.SimonJ. P.2000Molecular forms and kinetic properties of phosphoenolpyruvate carboxylase from barnyard grass (Echinochloa crus-galli(L.) Beauv.: Poaceae) Canadian Periodical of Botany78619-628.
27.HamiltonEast. W.HeckathornDue south. A.JoshiP.WangD.BaruaD.2008Interactive Effects of Elevated CO2 and Growth Temperature on the Tolerance of Photosynthesis to Acute Heat Stress in C3 and C4 Species.Journal of Integrative Constitute Biologylxi): 1375-1387.
28.HatchM. D.1987C4 photosynthesis: a unique blend of modified biochemistry, beefcake and ultrastructure.Biochimica et Biophysica Acta89581-106.
29.Hatch1000. D.1992Civ photosynthesis: an unlikely process full of surprises.Institute Cell Physiology33333-342.
30.HattersleyP. W.198213 values in Civ types in grasses.Australian Journal of Establish Physiology.ix139-154.
31.HeckathornDue south. A.DownsC. A.SharkeyT. D.ColemanJ. Due south.1998The modest, methionine-rich chloroplast heat-shock protein protects photosystem II electron transport during heat stress.Plant Physiology116439-444.
32.HeckathornSouthward. A.RyanS. L.BaylisJ. A.WangJ. A.HamiltonEast. West.CundiffL.et al.2002In vivo evidence from an Agrostis stolonifera choice genotype that chloroplast small rut-shock proteins tin can protect photosystem 2 during heat stress. Functional Plant Biology29933-944.
33.HuxmanT. East.HamerlynckE. P.LoikM. E.SmithSouthward. D.1998Gas substitution and chlorophyll fluorescence responses of three southwesternYuccaspecies to elevated CO2 and high temperature. Plant Prison cell and Environment211275-1283.
34.Intergovernmental Panel on Climate Modify, IPCC2001Climate change 2001: the scientific basis. In:Contribution of Working Group I to the 3rd Assessment Report of the Intergovernmental Panel on Climate Change Summary for Policymakers. Houghton, J.T.; Ding, Y.; Griggs, D.J.; Noguer, Yard.; van der Linden, P.J.; Dai, X.; Maskell, K. & Johnson, C.A., (Eds.) Cambridge University Press, Cambridge, U.k.,881
35.Intergovernmental Panel on Climate Change, IPCC2007Climatic change 2007: synthesis study. In: Pachauri, R.K. & Reisinger, A. (Eds.)Contribution of Working Groups I, II and 3 to the Fourth Assessment Written report of the Intergovernmental Console on Climate Modify. IPCC, Geneva.104
36.KirschbaumThou. U. F.2010Does Enhanced Photosynthesis Heighten Growth? Lessons Learned from CO2 Enrichment Studies.Plant Physiology155117-124.,
37.KrallJ. P.EdwardsG. Eastward.1993PEP carboxylases from 2 C4 species of Panicum with markedly different susceptibilities to cold inactivation. Found and Jail cell Physiology341-11.
38.KubienD. S.von CammererS.FurbankR. T.SageR. F.2003Civ photosynthesis at depression temperature. A study using transgenic plants with reduced amounts of Rubisco.Institute Physiology1321577-1585.
39.LaraG. V.AndreoC. S.2005Photosynthesis in not typical Civ species In: Pessarakli, M. (Ed.),Handbook of Photosynthesis: 2nd Edition"CRC press, Taylor & Francis Group. Boca Ratón, FL, United states of america,391421.
twoscore.LaraK. V.CasatiP.AndreoC. S.2002CO2 concentration mechanisms in Egeria densa, a submersed aquatic species. Physiologia Plantarum115487-495.
41.LawlorD. W.KontturiYard.ImmatureA. T.1989Photosynthesis past flag leaves of wheat in relation to protein, ribulose bisphosphate carboxylase activity and nitrogen supply. Journal of Experimental Botanyxl43-52.
42.LeakeyA. D. B.AinsworthE. A.BernacchiC. J.RogersA.LongS. P.OrtD. R.2009Elevated CO2 effects on institute carbon, nitrogen and h2o relations: six important lessons from Confront.Periodical of Experimental Botanylx2859-2876.
43.LeakeyA. D. B.UribelarreaM.AinsworthEast. A.NaiduSouth. L.RogersA.OrtD. R.LongDue south. P.2006Photosynthesis, productivity, and yield of maize are non affected by open-air acme of COii concentration in the absenteeism of drought.Plant Physiology140779790.
44.LongSouthward. P.1991Modification of the response of photosynthetic productivity to rise temperature by atmospheric CO2 concentrations: has its importance been underestimated?Plant Jail cell and Surroundxiv729-739.
45.LongS. P.1999Environmental responses. In: Sage, R.F. & Monson, R.K. (Eds.).C4 establish biology.Academic Printing, San Diego, United states,215249.
46.LongSouthward. P.AinsworthEast. A.RogersA.OrtD. R.2004Rise atmospheric carbon dioxide: Plants Face up the time to come.Almanac Reviews in Constitute Biology55591-628.
47.LongS. P.ZhuX. G.NaiduS. L.OrtD. R.2006Can improvement in photosynthesis increase crop yields?Found Cell and Environment29315-330.
48.Ludwig50. J.CanvinD. T.1971Charge per unit of photorespiration during photosynthesis and relationship of substrate of light respiration to products of photosynthesis in sunflower leaves.Plant Physiology48712719.
49.MatsubaK.ImaizumiDue north.KaneloS.AmejimaM.OhsugiR.1997Photosynthetic responses to temperature of phosphoenolpyruvate carboxykinase type Civ species differing in cold sensitivity.Establish Prison cell and Surroundings20268-274.
50.MorisonJ. I. L.LawlorD. W.1999Interactions between increasing COtwo concentration and temperature on constitute growth.Plant Cell and Surroundings22659-682.
51.PiedadeM. T. F.JunkWestward. J.LongS. P.1991The productivity of the C4 grassEchinochloa polystachyaon the Amazon floodplain. Ecology721456-1463.
52.PittermannJ.SageR. F.2000Photosynthetic performance at low temperature of Bouteloua gracilis Lag., a loftier-altitude C4 grass from the Rocky Mountains, USA.Constitute, Cell and Environs23811823.
53.PooterH.NavasThou. L.2003Plant growth and competition at elevated CO2: on winners, losers and functional groups.New Phytologist157175-198.
54.PortisA. R.JrParryM. A. J.2007Discoveries in Rubisco (ribulose 1,5-bisphosphate carboxylase/oxygenase): a historical perspective.Photosynthesis Inquiry94121-143.
55.ReddyA. R.RasineniG. Grand.RaghavendraA. S.2010The impact of global elevated CO2 concentration on photosynthesis and plant productivity.Current Science9946-57.
56.ReichP. B.TilmanD.CraineJ.EllsworthD.TjoelkerM.MarkThou.et al.2001Do species and functional groups differ in acquisition and use of C, North and h2o under varying atmospheric COii and N availability regimes? A field test with 16 grassland species.New Phytologist150435-448.
57.RipleyB. S.AbrahamT. I.OsborneC. P.2008Consequences of C4 photosynthesis for the partitioning of growth: a test using C3 and Cfour subspecies of Alloteropsis semialata under nitrogen-limitation. Journal of Experimental Phytology5971705-1714
58.RodenJ. South.BallM. C.1996Growth and photosynthesis of two eucalypt species during loftier temperature stress nether ambient and elevated [CO2].Global Change Biological science2115-128.
59.SageR. F.2002How terrestrial organisms sense, betoken and respond to carbon dioxide.Integrative Computational Biology42469-480.6,7. T
60.SageR. F.Mc KownA. D.2006Is Cfour photosynthesis less phenotypically plastic than C3 photosynthesis?Journal of Experimental Phytology572303-317.
61.SageR. F.MonsonR. Grand.1999C4 constitute biology.Academic Press, San Diego.
62.SageR. F.PearcyR. W.2000The physiological ecology of C4 photosynthesis. Kluwer, Dordretch, The netherlands.
63.SageR. F.KubienD. Due south.2003Quo vadis C4? An ecophysiological perspective on global change and the futurity of C4 plants. Photosynthesis Research772-3, 209-225.
64.SageR. F.1994Acclimation of photosynthesis to increasing atmospheric CO2-the gas-exchange perspective.Photosynthesis Research39351-368.
65.SageR. F.2002Variation in the k(cat) of Rubisco in C3 and C4 plants and some implications for photosynthetic performance at loftier and low temperature.Periodical of Experimental Phytology53609-620.
66.SageR. F.KocacinarF.KubienD. Due south.2011Civ photosynthesis and temperature. In: Raghavendra, A.S. & Sage, R.F. (Eds.), Cfour Photosynthesis and Related CO2 Concentrating Mechanisms,Springer Scientific discipline+Business Media B.V., Dordrecht, The Netherlands,161195.
67.SageR. F.PearcyR. W.SeemannJ. R.1987The nitrogen use efficiency of C3 and C4 plants. Three. Foliage nitrogen effects on the activeness of carboxylating enzymes inChenopodium album(L.) and Amaranthus retroflexus (50.). Plant Physiology85355-359.
68.SageR. F.WayD. A.KubienD. South.2008Rubisco, Rubisco activase, and global climatic change.Journal of Experimental Botany591581-1595.
69.SaliendraN. Z.MeinzerF. C.PerryM.ThomM.1996Associations between partitioning of carboxylase activity and bundle sheath leakiness to CO2, carbon isotope discrimination, photosynthesis, and growth in sugarcane. Journal of Experimental Botany47907914.
70.SantariusThousand. A.1975The protective outcome of sugars on chloroplast membranes during temperature and water stress and its relationship to frost, desiccation and oestrus resistance.Periodical of Thermal. Biology1101-107.
71.SchiermeierQ.2008H2o: A long dry summer.Nature452270-273.
72.StilingP.CornelissenT.2007How does elevated carbon dioxide (CO2) affect found-herbivore interactions? A field experiment and metaanalysis of CO2-mediated changes on plant chemistry and herbivore performance.Global Alter Biologythirteen1823-1842.
73.TaubD. R.SeemannJ. R.ColemanJ. S.2000Growth in elevated CO2 protects photosynthesis against loftier-temperature damage.Plant Cell and Environment23649-656.
74.ThomasC. D.CameronA.GreenR. E.BakkenesChiliad.BeaumontL. J.CollinghamY. C.et al.2004Extinction risk from climate modify.Nature427145-158.
75.WallG. W.BrooksT. J.AdamR.et al.2001Elevated atmospheric CO2 improved sorghum plant water status past ameliorating the adverse effects of drought.New Phytologist152231-248.
76.WangD.HeckathornSouth. A.BaruaD.JoshiP.HamiltonEast. W.La CroixJ.2008Effects of elevated CO2 on the tolerance of photosynthesis to acute heat stress in C3, C4, and CAM species.American Journal of Botany95165-176.
77.WangTen.2007Furnishings of species richness and elevated carbon dioxide on biomass aggregating: a synthesis using meta-analysis.Oecologia152595-605.
78.WardJ. Grand.TissueD. T.ThomasR. B.StrainB. R.1999Comparative responses of model C3 and C4 plants to drought in low and elevated COtwo.Global Alter Biological sciencev857-867.
79.WeisE.BerryJ. A.1988Plants and high temperature stress, In: Long; Southward.P. & Woodward F.I. (Eds.),Plants and Temperature, The Visitor of Biologists Ltd., Cambridge,329346.
80.ZiskaL. H.SicherR. C.BunceJ. A.1999The bear upon of elevated carbon dioxide on the growth and gas exchange of three Cfour species differing in CO2 leak rates.Physiologia Plantarum10574-80.
Written By
María Valeria Lara and Carlos Santiago Andreo
Submitted: December 16th, 2010Reviewed: April 29th, 2011Published: September 22nd, 2011





0 Response to "In Which Type Of Environment Are Plants Most Likely To Use A C4 Pathway Of Carbon Fixation?"
Post a Comment